The new JVP
JVP is the Journal of Vertebrate Paleontology. There are lots of paleo journals, but I think it is fair to say that JVP is one of the flagships, at least for people working on vertebrates. It's always packed with data, but some issues are just more interesting than others. Sometimes it's all partial fish scales from the Apathian of South New Brunswick and heavily eroded rodent teeth from Devil's Outhouse National Monument.
But the current issue is a gold mine.
Witzmann (2006) reports on patterns of skeletal development in a Permo-Carboniferous temnospondyl (a pre-dinosaurian giant salamander relative). Hold on, there. It's more interesting than it may sound. These temnospondyls were filling the ecospace occupied by crocs and aquatic turtles and true salamanders today. Most of them went through a metamorphosis like living amphibians. Some have been shown to have two adult morphs, one metamorphosed and one neotenic, just like the living salamander Ambystoma with its axolotl form. Witzman's figures ROCK. Figs 5 and 6 show developmental patterns for several temnospondyls and other ancient salamanderish critters and for Ambystoma. The patterns are ossification sequences for various bones in the body, arranged into vertical stacks representing relative time, with lines connecting equivalent steps in adjacent columns so that you can see where the developmental deck has been heterochronically shuffled. Very cool.
UPDATE Florian gave me permission to post his figure. Check it out.
Trueb and Baez (2006)--yes, that Trueb, for you herp-savvy folks--redescribe some itty bitty pipoid frogs from the Cretaceous of Israel. The frogs are not terribly mind-blowing as frogs, unless you're really invested in pipoid phylogeny. I just like learning about all the other critters that were running around during the Cretaceous. It pleases me to think that if I was shot back in a time machine I might have just as good a chance at identifying some of the frogs and lizards as I would at the dinosaurs.
Gower and Nesbitt (2006) describe the braincase of Arizonasaurus and show that "rauisuchians", a group of terrestrial, predatory, pre-dinosaurian croc relatives, are not monophyletic. Some are closer to crocs than others. Arizonasaurus and its allies, which include such weirdos as Lotosaurus, are pretty far down the tree from crocs. Which is fine. If you saw one you would not think "terrestrial croc relative". You'd think, Why is that theropod trucking around on all fours, and why does it have a sail on it's back?" The bird-line archosaurs--pterosaurs and dinos, mostly--have been hogging the limelight for the past, well, forever, but that may be about to change. Effigia and Arizonsaurus are just the beginning.
Here's a picture of Lotosaurus so that you may grok its prehistoric weirdness. The sailback is independently evolved in this group--it has nothing to do with Dimetrodon and Edaphosaurus, which are closer to mammals, or Ouranosaurus, Acrocanthosaurus, and Spinosaurus, which are all closer to birds. Sails were just popular back in the day.

Next up, a Berkeley-MSU paper: Mark Goodwin, Bill Clemens, Jack Horner, and Kevin Padian (2006) on a new dinky skull from a baby Triceratops. This one made the cover. The little skull is adorable, about 2/3 complete, and about a foot long. It already has little brow horns and a scalloped frill. Goodwin et al. take a swipe at the prevailing view that ceratopsians horns and frills were primarily for sexual display. Sexual display structures are usually sexually dimorphic--that's the whole idea, that an individual can tell the boys from the girls. Trouble is, demonstrating sexual dimorphism is not easy. It requires a large enough sample size that sex-related differences can be parsed out from differences related to growth and simple within-population variation. So far the only ceratopsian that meets those standards is Protoceratops. That doesn't mean that the horns and frills of more derived ceratopsians weren't used in sexual display, just that convincing evidence has not been presented for it. Goodwin et al. favor the hypothesis that the fancy headgear was mainly for species recognition: it varies a lot between closely related taxa, but not in a linear fashion that would indicate improving function. It is probably dumb to deny that horns would be useful in fending off tyrannosaurs, but it is worth remembering that centrosaurines started out with horns and then traded them in for big bosses of rough bone on their noses--not what you'd expect if horns were absolutely necessary for their survival. Finding decently developed horncores and shield morphology in a little baby is also more consistent with the species recognition hypothesis than the sexual display hypothesis, although Goodwin et al. allow that the three functions--species recognition, defense, and sexual display--are not mutually exclusive and could have all contributed to the weird heads of ceratopsians.
I don't have a picture of the little Triceratops, but here's a picture of its relative Pentaceratops. This unusually large individual from New Mexico has a skull 10.5 feet long, possibly the longest of any terrestrial vertebrate (unless there are some new Torosaurus specimens I don't know about). You can see that the hindlimb bones have been smooshed flat and parts of the skeleton have been reconstructed. The brow horns on that thing are a full meter long, and they would have been longer with the keratin sheath on (like the horns of cattle). I swiped the photo from http://sill-www.army.mil/pao/Photos/030626/pages/Pentaceratops.htm.

Conrad (2006) describes a new lizard from the Eocene (~50 million years ago) of Wyoming. The lizard is strikingly similar to the living Chinese crocodile lizard, which is cool, because up until now that thing had no fossil record at all. It's closest relatives are from the Cretaceous at least, which means that like the coelocanth it has survived for at least 65 million years without leaving any fossils--until now. Conrad plugged the crocodile lizard and its new Eocene relative into a phylogenetic analysis and found that they branched off from the monitor lizard lineage almost 100 million years ago. So as the last brachiosaurs in North America were going extinct, and as our ancestors were getting used to these newfangled placenta things, the first crocodile lizards were setting sail on their long, lonely voyage through time. And they're still around. Cretaceous survivors rock.
DeBlieux et al. (2006) describe an internal mandibular chamber in Eocene hyraxes that shows sexual dimorphism (it was a big issue for Eocene critters and testing sexual dimorphism). What's all that mean? Hyraxes are also known colloquially as coneys. They are little marmot-looking critters that you may have seen crawling around in rock piles at the zoo. Oddly enough, their closest living relatives are elephants and manatees. No shit. I should point out that the living hyraxes are little marmot-looking things, but for much of the Age of Mammals there were bigass hyracoids that looked tapirs and rhinos. In fact, they seem to have been replaced ecologically by tapirs and rhinos. These Eocene critters have big hollow chambers inside their lower jaws. Holes in bones have to be filled by something, and there are only so many possibilities. Basically it comes down to muscles, salivary glands, and air sacs. The muscular hypothesis is pretty easy to rule out. Muscles leave scars on bones, and the muscle scars on the hyrax jaws do not extend into the jaw cavities. Salivary glands are found in adjacent positions in living hyraxes, but in general glands are not found within bony cavities. Also, there's no obvious reason why one sex would need a lot more saliva than the other. (Yeah, I know how that sounds. If I can leave it alone, so can you.) What about air sacs? The case actually looks pretty good. The bony signature is similar to what you get with air-filled bones, and lots of mammals, including hyraxes, have air-filled pouches that develop from the larynx, trachea, or eustachian tubes. DeBlieux et al. posit that the jaw cavities were filled with air and functioned as resonators for making calls, like the inflated upper jaws of living pacas.
Several things impress me about this paper. The first is the careful way in which the authors evaluate the competing hypotheses. They admit that they can't rule out the glandular hypothesis, it just has a lot less going for it than the air sac hypothesis. And they are also careful to separate the anatomical hypothesis (the cavity contained an air sac) from the functional one (the air sac was used as a resonator for making calls). Finally, their review of the literature on air sacs in mammals is a godsend to anyone who is interested in this arcane topic. I didn't even know that horses had guttural sacs, but evidently there was a paper on their function in Nature in 2000. This is exactly the kind of thing I have in mind when I write about how much enjoyment I get out of being surprised by the living world.
One more and then I'm packing it in. Henderson and Peterson (2006) describe a neck vertebra of an azhdarchid pterosaur from the Hell Creek Formation. Hell Creek is where the first skeletons of Tyrannosaurus were found. In fact, this pterosaur vert came out of a plaster jacket thrown around some tyrannosaur bones! Azhdarchids are the group that includes Quetzalcoatlus, Arambourgenia, and Hatzegopteryx, all of which are contenders for the largest flying animals of all time. The find is cool for several reasons. It's the first diagnostic piece of pterosaur to be published from the whole Hell Creek Formation. Hell Creek is not good country for finding pterosaurs. Like most non-lagoon deposits, it's just too coarse to preserve immensely fragile pterosaur bones, which on average were even more lightly built than the bones of birds. It's not that the pterosaurs weren't there, just that they very rarely made it into the fossil record. The vert is also wicked slender. Azhdarchids are weird; they've got just a double handful of very, very long neck vertebrae, which would not have made for a very flexible neck (or a very strong one, you would think). You can grok some azhdarchid weirdness and read up on what they were doing with those wicked necks here.
Now, I know from long-ass necks. Sauroposeidon (which, yes, I am obligated to mention in every single post--deal with it) was no slouch in the long neck department. Its most slender vertebrae have a length to diameter ratio of about 6.5. There are maybe one or two sauropods out there with vertebrae as extreme. Mamenchisaurus youngi and the newly-named Erketu are the only ones that come to mind. But these 'pods were pikers compared to azhdarchids. The vertebra described by Henderson and Peterson is 37 cm long. It's crushed, but the ends aren't in too bad of shape. The diameter at the posterior end was probably no more than 2.5 cm, which would give it a length to diameter ratio of, gulp, 15.
I know you're dying to see Sauroposeidon get its 50-ton ass handed to it by a pterosaur the mass of a turkey, so here you go.
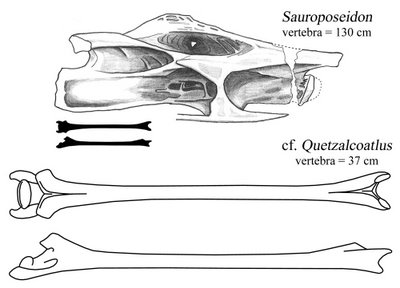 The pterosaur vert I traced from Henderson and Peterson (2006:fig. 3). Both verts are shown with the front end to the right. The upper pterosaur drawing is the vert from above, and the lower is from the right side. It is worth remembering that not only is that vertebra more slender than most limb bones, but it was also about 80-90% air (same goes for Sauroposeidon, come to think of it). If that doesn't blow your mind, then I pronounce your mind unblowable. And I pity you.
The pterosaur vert I traced from Henderson and Peterson (2006:fig. 3). Both verts are shown with the front end to the right. The upper pterosaur drawing is the vert from above, and the lower is from the right side. It is worth remembering that not only is that vertebra more slender than most limb bones, but it was also about 80-90% air (same goes for Sauroposeidon, come to think of it). If that doesn't blow your mind, then I pronounce your mind unblowable. And I pity you.
That's just the tip of the iceberg. Those are just seven papers out of 25 in this issue. I didn't mention the new fish, or the other new amphibians, or the new ichthyosaur, or two other new lizards, or the new jumping mouse, or the new pangolin, or the new whale. And, shockingly, I didn't discuss the one sauropod paper, Jerry Harris's description of the skull material of Suuwassea.
I'll probably never do this again. It has taken up half the day. But I was jazzed by all the new hotness, and I just had to share it.
Here are those refs.
-------------------------------------------------
DeBlieux, D.D., et al. 2006. Sexual dimorphism of the mandibular chamber in Fayum Pliohyracidae (Mammalia). JVP 26(1):160-169.
Conrad, J. 2006. An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, U.S.A. JVP 26(1):113-126.
Goodwin, M.B., et al. 2006. The smallest known Triceratops skull: new observations on ceratopsid cranial anatomy and ontogeny. JVP 26(1):103-112.
Gower, D.J., and Nesbitt, S.J. 2006. The braincase of Arizonasaurus babbitti--further evidence for the non-monophyly of 'rauisuchian' archosaurs. JVP 26(1):79-87.
Harris, J.D. 2006. Cranial osteology of Suuwassea emilieae (Sauropoda: Diplodocoidea: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. JVP 26(1):88-102.
Henderson, M.D., and Peterson, J.E. 2006. An azhdarchid pterosaur cervical vertebra from the Hell Creek Formation (Maastrichtian) of southeastern Montana. JVP 26(1):192-195.
Trueb, L., and Baez, A.M. 2006. Revision of the Early Cretaceous Cordicephalus from Israel and an assessment of its relationships among pipoid frogs. JVP 26(1):44-59.
Witzmann, F. 2006. Developmental patterns and ossification sequence in the Permo-Carboniferous temnospondyl Archegosaurus decheni (Saar-Nahe Basin, Germany). JVP 26(1):7-17.
But the current issue is a gold mine.
Witzmann (2006) reports on patterns of skeletal development in a Permo-Carboniferous temnospondyl (a pre-dinosaurian giant salamander relative). Hold on, there. It's more interesting than it may sound. These temnospondyls were filling the ecospace occupied by crocs and aquatic turtles and true salamanders today. Most of them went through a metamorphosis like living amphibians. Some have been shown to have two adult morphs, one metamorphosed and one neotenic, just like the living salamander Ambystoma with its axolotl form. Witzman's figures ROCK. Figs 5 and 6 show developmental patterns for several temnospondyls and other ancient salamanderish critters and for Ambystoma. The patterns are ossification sequences for various bones in the body, arranged into vertical stacks representing relative time, with lines connecting equivalent steps in adjacent columns so that you can see where the developmental deck has been heterochronically shuffled. Very cool.
UPDATE Florian gave me permission to post his figure. Check it out.
Trueb and Baez (2006)--yes, that Trueb, for you herp-savvy folks--redescribe some itty bitty pipoid frogs from the Cretaceous of Israel. The frogs are not terribly mind-blowing as frogs, unless you're really invested in pipoid phylogeny. I just like learning about all the other critters that were running around during the Cretaceous. It pleases me to think that if I was shot back in a time machine I might have just as good a chance at identifying some of the frogs and lizards as I would at the dinosaurs.
Gower and Nesbitt (2006) describe the braincase of Arizonasaurus and show that "rauisuchians", a group of terrestrial, predatory, pre-dinosaurian croc relatives, are not monophyletic. Some are closer to crocs than others. Arizonasaurus and its allies, which include such weirdos as Lotosaurus, are pretty far down the tree from crocs. Which is fine. If you saw one you would not think "terrestrial croc relative". You'd think, Why is that theropod trucking around on all fours, and why does it have a sail on it's back?" The bird-line archosaurs--pterosaurs and dinos, mostly--have been hogging the limelight for the past, well, forever, but that may be about to change. Effigia and Arizonsaurus are just the beginning.
Here's a picture of Lotosaurus so that you may grok its prehistoric weirdness. The sailback is independently evolved in this group--it has nothing to do with Dimetrodon and Edaphosaurus, which are closer to mammals, or Ouranosaurus, Acrocanthosaurus, and Spinosaurus, which are all closer to birds. Sails were just popular back in the day.

Next up, a Berkeley-MSU paper: Mark Goodwin, Bill Clemens, Jack Horner, and Kevin Padian (2006) on a new dinky skull from a baby Triceratops. This one made the cover. The little skull is adorable, about 2/3 complete, and about a foot long. It already has little brow horns and a scalloped frill. Goodwin et al. take a swipe at the prevailing view that ceratopsians horns and frills were primarily for sexual display. Sexual display structures are usually sexually dimorphic--that's the whole idea, that an individual can tell the boys from the girls. Trouble is, demonstrating sexual dimorphism is not easy. It requires a large enough sample size that sex-related differences can be parsed out from differences related to growth and simple within-population variation. So far the only ceratopsian that meets those standards is Protoceratops. That doesn't mean that the horns and frills of more derived ceratopsians weren't used in sexual display, just that convincing evidence has not been presented for it. Goodwin et al. favor the hypothesis that the fancy headgear was mainly for species recognition: it varies a lot between closely related taxa, but not in a linear fashion that would indicate improving function. It is probably dumb to deny that horns would be useful in fending off tyrannosaurs, but it is worth remembering that centrosaurines started out with horns and then traded them in for big bosses of rough bone on their noses--not what you'd expect if horns were absolutely necessary for their survival. Finding decently developed horncores and shield morphology in a little baby is also more consistent with the species recognition hypothesis than the sexual display hypothesis, although Goodwin et al. allow that the three functions--species recognition, defense, and sexual display--are not mutually exclusive and could have all contributed to the weird heads of ceratopsians.
I don't have a picture of the little Triceratops, but here's a picture of its relative Pentaceratops. This unusually large individual from New Mexico has a skull 10.5 feet long, possibly the longest of any terrestrial vertebrate (unless there are some new Torosaurus specimens I don't know about). You can see that the hindlimb bones have been smooshed flat and parts of the skeleton have been reconstructed. The brow horns on that thing are a full meter long, and they would have been longer with the keratin sheath on (like the horns of cattle). I swiped the photo from http://sill-www.army.mil/pao/Photos/030626/pages/Pentaceratops.htm.

Conrad (2006) describes a new lizard from the Eocene (~50 million years ago) of Wyoming. The lizard is strikingly similar to the living Chinese crocodile lizard, which is cool, because up until now that thing had no fossil record at all. It's closest relatives are from the Cretaceous at least, which means that like the coelocanth it has survived for at least 65 million years without leaving any fossils--until now. Conrad plugged the crocodile lizard and its new Eocene relative into a phylogenetic analysis and found that they branched off from the monitor lizard lineage almost 100 million years ago. So as the last brachiosaurs in North America were going extinct, and as our ancestors were getting used to these newfangled placenta things, the first crocodile lizards were setting sail on their long, lonely voyage through time. And they're still around. Cretaceous survivors rock.
DeBlieux et al. (2006) describe an internal mandibular chamber in Eocene hyraxes that shows sexual dimorphism (it was a big issue for Eocene critters and testing sexual dimorphism). What's all that mean? Hyraxes are also known colloquially as coneys. They are little marmot-looking critters that you may have seen crawling around in rock piles at the zoo. Oddly enough, their closest living relatives are elephants and manatees. No shit. I should point out that the living hyraxes are little marmot-looking things, but for much of the Age of Mammals there were bigass hyracoids that looked tapirs and rhinos. In fact, they seem to have been replaced ecologically by tapirs and rhinos. These Eocene critters have big hollow chambers inside their lower jaws. Holes in bones have to be filled by something, and there are only so many possibilities. Basically it comes down to muscles, salivary glands, and air sacs. The muscular hypothesis is pretty easy to rule out. Muscles leave scars on bones, and the muscle scars on the hyrax jaws do not extend into the jaw cavities. Salivary glands are found in adjacent positions in living hyraxes, but in general glands are not found within bony cavities. Also, there's no obvious reason why one sex would need a lot more saliva than the other. (Yeah, I know how that sounds. If I can leave it alone, so can you.) What about air sacs? The case actually looks pretty good. The bony signature is similar to what you get with air-filled bones, and lots of mammals, including hyraxes, have air-filled pouches that develop from the larynx, trachea, or eustachian tubes. DeBlieux et al. posit that the jaw cavities were filled with air and functioned as resonators for making calls, like the inflated upper jaws of living pacas.
Several things impress me about this paper. The first is the careful way in which the authors evaluate the competing hypotheses. They admit that they can't rule out the glandular hypothesis, it just has a lot less going for it than the air sac hypothesis. And they are also careful to separate the anatomical hypothesis (the cavity contained an air sac) from the functional one (the air sac was used as a resonator for making calls). Finally, their review of the literature on air sacs in mammals is a godsend to anyone who is interested in this arcane topic. I didn't even know that horses had guttural sacs, but evidently there was a paper on their function in Nature in 2000. This is exactly the kind of thing I have in mind when I write about how much enjoyment I get out of being surprised by the living world.
One more and then I'm packing it in. Henderson and Peterson (2006) describe a neck vertebra of an azhdarchid pterosaur from the Hell Creek Formation. Hell Creek is where the first skeletons of Tyrannosaurus were found. In fact, this pterosaur vert came out of a plaster jacket thrown around some tyrannosaur bones! Azhdarchids are the group that includes Quetzalcoatlus, Arambourgenia, and Hatzegopteryx, all of which are contenders for the largest flying animals of all time. The find is cool for several reasons. It's the first diagnostic piece of pterosaur to be published from the whole Hell Creek Formation. Hell Creek is not good country for finding pterosaurs. Like most non-lagoon deposits, it's just too coarse to preserve immensely fragile pterosaur bones, which on average were even more lightly built than the bones of birds. It's not that the pterosaurs weren't there, just that they very rarely made it into the fossil record. The vert is also wicked slender. Azhdarchids are weird; they've got just a double handful of very, very long neck vertebrae, which would not have made for a very flexible neck (or a very strong one, you would think). You can grok some azhdarchid weirdness and read up on what they were doing with those wicked necks here.
Now, I know from long-ass necks. Sauroposeidon (which, yes, I am obligated to mention in every single post--deal with it) was no slouch in the long neck department. Its most slender vertebrae have a length to diameter ratio of about 6.5. There are maybe one or two sauropods out there with vertebrae as extreme. Mamenchisaurus youngi and the newly-named Erketu are the only ones that come to mind. But these 'pods were pikers compared to azhdarchids. The vertebra described by Henderson and Peterson is 37 cm long. It's crushed, but the ends aren't in too bad of shape. The diameter at the posterior end was probably no more than 2.5 cm, which would give it a length to diameter ratio of, gulp, 15.
I know you're dying to see Sauroposeidon get its 50-ton ass handed to it by a pterosaur the mass of a turkey, so here you go.
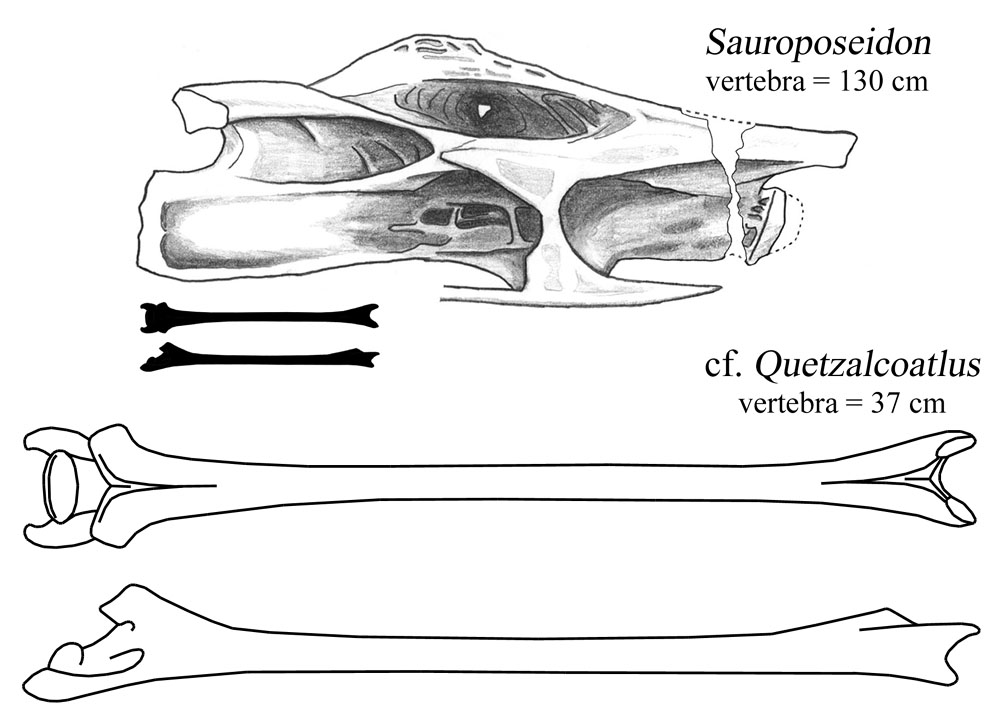 The pterosaur vert I traced from Henderson and Peterson (2006:fig. 3). Both verts are shown with the front end to the right. The upper pterosaur drawing is the vert from above, and the lower is from the right side. It is worth remembering that not only is that vertebra more slender than most limb bones, but it was also about 80-90% air (same goes for Sauroposeidon, come to think of it). If that doesn't blow your mind, then I pronounce your mind unblowable. And I pity you.
The pterosaur vert I traced from Henderson and Peterson (2006:fig. 3). Both verts are shown with the front end to the right. The upper pterosaur drawing is the vert from above, and the lower is from the right side. It is worth remembering that not only is that vertebra more slender than most limb bones, but it was also about 80-90% air (same goes for Sauroposeidon, come to think of it). If that doesn't blow your mind, then I pronounce your mind unblowable. And I pity you.That's just the tip of the iceberg. Those are just seven papers out of 25 in this issue. I didn't mention the new fish, or the other new amphibians, or the new ichthyosaur, or two other new lizards, or the new jumping mouse, or the new pangolin, or the new whale. And, shockingly, I didn't discuss the one sauropod paper, Jerry Harris's description of the skull material of Suuwassea.
I'll probably never do this again. It has taken up half the day. But I was jazzed by all the new hotness, and I just had to share it.
Here are those refs.
-------------------------------------------------
DeBlieux, D.D., et al. 2006. Sexual dimorphism of the mandibular chamber in Fayum Pliohyracidae (Mammalia). JVP 26(1):160-169.
Conrad, J. 2006. An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, U.S.A. JVP 26(1):113-126.
Goodwin, M.B., et al. 2006. The smallest known Triceratops skull: new observations on ceratopsid cranial anatomy and ontogeny. JVP 26(1):103-112.
Gower, D.J., and Nesbitt, S.J. 2006. The braincase of Arizonasaurus babbitti--further evidence for the non-monophyly of 'rauisuchian' archosaurs. JVP 26(1):79-87.
Harris, J.D. 2006. Cranial osteology of Suuwassea emilieae (Sauropoda: Diplodocoidea: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. JVP 26(1):88-102.
Henderson, M.D., and Peterson, J.E. 2006. An azhdarchid pterosaur cervical vertebra from the Hell Creek Formation (Maastrichtian) of southeastern Montana. JVP 26(1):192-195.
Trueb, L., and Baez, A.M. 2006. Revision of the Early Cretaceous Cordicephalus from Israel and an assessment of its relationships among pipoid frogs. JVP 26(1):44-59.
Witzmann, F. 2006. Developmental patterns and ossification sequence in the Permo-Carboniferous temnospondyl Archegosaurus decheni (Saar-Nahe Basin, Germany). JVP 26(1):7-17.
posted by Dr. Vector at 9:07 AM
![]()
![]()


0 Comments:
Post a Comment
<< Home